A&A 438, 1115-1122 (2005)
DOI: 10.1051/0004-6361:20042579
Time series study of EUV spicules observed by SUMER/SoHO
L. D. Xia 1,2 - M. D. Popescu 2,3 - J. G. Doyle2 - J. Giannikakis2,4
1 - CAS Key Laboratory of Basic Plasma Physics, School of Earth and Space Sciences,
Univ. of Science and Technology of China, Hefei, Anhui 230026, PR China
2 -
Armagh Observatory, College Hill, Armagh BT61 9DG, N. Ireland
3 -
Astronomical Institute of the Romanian Academy, 75212 Bucharest 28, Romania
4 -
Sect. of Astrophysics, Astronomy and Mechanics, Dept. of Physics, Univ. of Athens, Athens 15783, Greece
Received 21 December 2004 / Accepted 18 April 2005
Abstract
Here we study the dynamic properties of EUV spicules seen at the solar limb.
The selected data were obtained as time series in polar coronal holes by
SUMER/SoHO. The short exposure time and the almost fixed position of the spectrometer's
slit allow the analysis of spicule properties such as occurrence, lifetime and Doppler velocity.
Our data reveal that spicules occur repeatedly at the same location with a birth
rate of around 0.16/min as estimated at 10
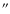 above the limb and a lifetime
ranging from 15 down to
above the limb and a lifetime
ranging from 15 down to 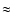 3 min. We are able to see some spicules showing a
process of "falling after rising'' indicated by the sudden change of the Doppler velocity sign.
A periodicity of 5 min is sometimes discernible in their occurrence.
Most spicules have a height between 10
and 20
above the limb. Some can stretch up to 40
;
these "long macro-spicules'' seem to
be comprised of a group of high spicules. Some of them have an obvious
periodicity in the radiance of 5 min.
3 min. We are able to see some spicules showing a
process of "falling after rising'' indicated by the sudden change of the Doppler velocity sign.
A periodicity of 5 min is sometimes discernible in their occurrence.
Most spicules have a height between 10
and 20
above the limb. Some can stretch up to 40
;
these "long macro-spicules'' seem to
be comprised of a group of high spicules. Some of them have an obvious
periodicity in the radiance of 5 min.
Key words: Sun: UV radiation - Sun: transition region
Solar spicules were seen on the limb more than a century ago (Secchi 1877; Roberts 1945).
They are rapidly evolving jet-like structures extending from the chromosphere upward into
the corona, and are probably the most common feature of the upper solar atmosphere. Spicules
were traditionally defined in the cool chromospheric lines such as H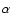 .
Reviews
summarizing H spicule properties have been given by Beckers (1972,1968)
and recently by Sterling (2000). Based on H observations (Beckers 1972,1968, and references
therein), typically, spicules have an average length ranging from 6500 km
to 9500 km, widths from several hundred to 2000 km and lifetimes from several minutes
to 15 min. They have an apparent upward velocity of about 25 km s-1.
After having reached the maximum height, spicules are found either falling
back into the chromosphere or fading out in the transition region (TR).
Spicules are reported to be slightly taller and oriented more nearly vertical in coronal holes (CHs),
and are somewhat more numerous at polar than at low latitudes.
Spicules are estimated to cover about 1% of the Sun's surface. Their
typical chromospheric density is 1011 cm-3. The upward mass flux
in spicules is estimated to be two orders of magnitude larger than that needed
to maintain the solar wind.
Yet, spicules are still among the most unresolved solar phenomena, as their physical
conditions are difficult to be clearly defined from observations due to their small size
and duration.
.
Reviews
summarizing H spicule properties have been given by Beckers (1972,1968)
and recently by Sterling (2000). Based on H observations (Beckers 1972,1968, and references
therein), typically, spicules have an average length ranging from 6500 km
to 9500 km, widths from several hundred to 2000 km and lifetimes from several minutes
to 15 min. They have an apparent upward velocity of about 25 km s-1.
After having reached the maximum height, spicules are found either falling
back into the chromosphere or fading out in the transition region (TR).
Spicules are reported to be slightly taller and oriented more nearly vertical in coronal holes (CHs),
and are somewhat more numerous at polar than at low latitudes.
Spicules are estimated to cover about 1% of the Sun's surface. Their
typical chromospheric density is 1011 cm-3. The upward mass flux
in spicules is estimated to be two orders of magnitude larger than that needed
to maintain the solar wind.
Yet, spicules are still among the most unresolved solar phenomena, as their physical
conditions are difficult to be clearly defined from observations due to their small size
and duration.
Since the launch of satellites, we are also able to observe spicule-like
structures above the limb in UV and EUV wavelengths. Bohlin et al. (1975) were the
first to clearly notice He II jets at the solar limb with the help of the XUV spectrograph on the Skylab mission. In their data, such jets were found to
construct a nearly continuous band over the polar caps, ranging from 5
to
over 60
in length (
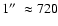 km on the Sun),
from 5
to 30
in width, and from 5 to over 40 min in lifetime. It should be noted that the term "macrospicules'' was then
used by Bohlin et al. to name the newly discovered EUV spicules in order to
distinguish them from the H spicules. In this context, the name "macrospicule''
is independent of the actual length of EUV spicules.
In the following sections, we use the term "long macro-spicule'' for these EUV spicules
that were occasionally observed as high as 30 000 km off-limb.
Note that, in the case of long H spicules, the term "H macrospicule'' is
used (see, e.g., Moore et al. 1977).
km on the Sun),
from 5
to 30
in width, and from 5 to over 40 min in lifetime. It should be noted that the term "macrospicules'' was then
used by Bohlin et al. to name the newly discovered EUV spicules in order to
distinguish them from the H spicules. In this context, the name "macrospicule''
is independent of the actual length of EUV spicules.
In the following sections, we use the term "long macro-spicule'' for these EUV spicules
that were occasionally observed as high as 30 000 km off-limb.
Note that, in the case of long H spicules, the term "H macrospicule'' is
used (see, e.g., Moore et al. 1977).
Table 1:
SUMER observations of temporal series in polar coronal
holes (PCHs).
From the Naval Research Laboratory (NRL) High Resolution Telescope and Spectrograph
(HRTS) and the Harvard experiment on Skylab, more information about
EUV spicules emerged (see, e.g., Dere et al. 1983; Karovska & Habbal 1994; Withbroe 1983; Dere et al. 1989).
Nevertheless, EUV spicules are more difficult to observe than the H ones, not only because the spatial resolution needed to study spicules is
more difficult to achieve with EUV instruments in space, but also because
they are much harder to detect against the bright TR background.
The best spectral and spatial resolution currently available in the EUV for solar studies comes from SUMER (Solar Ultraviolet Measurements of
Emitted Radiation), a spectrometer on SoHO (the Solar and Heliospheric
Observatory). Its spatial resolution of 1
,
together with a high
spectral resolution and a time cadence of
a few seconds, makes it a good instrument for resolving small-scale and
rapidly evolving structures such as spicules.
According to SUMER observations (Wilhelm 2000; Wilhelm et al. 2000; Budnik et al. 1998),
EUV spicules have a length comparable to H spicules. They have
widths of about 1-2
when seen at chromospheric temperatures, and are
wider in TR lines. Their enhanced radiance structures can be distinguishable
up to a temperature of about
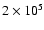 K. The line-of-sight (LOS)
velocity derived from the Doppler shift has average values of
K. The line-of-sight (LOS)
velocity derived from the Doppler shift has average values of 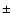 30 km s-1.
30 km s-1.
Spicule studies are still in their infancy, and many questions about their
properties are not yet answered, not to mention the efforts which are being
made for finding the mechanism that is producing them. There is a strong
debate as to whether they are due to propagating waves or to magnetic reconnection.
Another one of the most controversial issues is the connection between
H and EUV spicules. Although both features look like thin elongated
jets, a direct comparison is rather difficult (see, e.g., discussion in Sterling 2000). Because the density of EUV spicules measured at TR temperatures
is much lower and their width is much broader than those of H spicules,
it is suggested that, if these two structures belong to the same phenomenon, the
EUV spicules are likely the hot sheath of the cool H spicules, which
constructs a transition between the hotter corona and the cooler
material from below (Withbroe 1983).
In the attempt to elucidate more of the spicule characteristics,
in this contribution we study the dynamic properties of EUV spicules. The
data selected for this study were obtained as time series in polar CHs by
SUMER. The short exposure time (compared with the lifetime of spicules)
and the almost fixed position (due to a reduced effect of the solar rotation
at the poles) allow us to analyse the spicule occurrence, lifetime, LOS
velocity and other characteristics derived from the good spectral information
of our data. The temporal evolution of solar EUV spicules has never previously
been seen for such long observing sequences or with such clarity as reported
here. A colour version of the figures from this paper is given on-line.
![\begin{figure}
\par\includegraphics[width=8.8cm,clip]{2579fig1.ps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/Timg13.gif) |
Figure 1:
The two figures illustrate the solar poles in the light emitted by the
Fe XII 195 Å line (
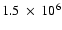 K), taken by EIT/SoHO.
The vertical white lines show the fixed position of the SUMER slit
during the observations for PCH 1 and PCH 2a.
The third dataset, PCH 2b, is taken in the same southern hole, four days before the PCH 2a data. K), taken by EIT/SoHO.
The vertical white lines show the fixed position of the SUMER slit
during the observations for PCH 1 and PCH 2a.
The third dataset, PCH 2b, is taken in the same southern hole, four days before the PCH 2a data. |
| Open with DEXTER |
![\begin{figure}
\par\includegraphics[width=16.8cm,clip]{2579fig2.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/Timg14.gif) |
Figure 2:
The temporal evolution of the northern coronal hole (PCH 1) seen in the N IV 765 Å ( left) and
Ne VIII 770 Å ( right) lines, with a cadence of 30 s. Spicule structures are clearly seen in the
TR line N IV (140 000 K), but not in the higher temperature line Ne VIII (630 000 K). At the top of the figures we show the radiance varying
with time for both lines at the height of 10
above the
limb (indicated, in the 2D plots, by the dotted black line).
The continuous black line represents the N IV limb. |
| Open with DEXTER |
The data selected for this study were obtained as time series in two polar
coronal holes (PCHs, see Table 1) by the SUMER spectrometer
(Wilhelm et al. 1997; Lemaire et al. 1997; Wilhelm et al. 1995). During the observation, the SUMER
slit was fixed (without scanning) at position x=0
,
y=909.75
for PCH 1,
x=0
,
y=-950.25
for PCH 2a, and x=0
,
y=-984.75
for PCH 2b. The two CHs are shown in Fig. 1,
where the EIT (Extreme Ultraviolet Imaging Telescope) 195 Å images are plotted overlaid
with the position of the SUMER slit. The two PCHs (dark regions) can be clearly distinguished
in the images.
The slit width (1
for PCH 1 and PCH 2a; 0.3
for PCH 2b)
determine the spatial resolution along the x-direction, while the resolution
element along the slit in the y-direction (north-south; positive towards
north) is given by the pixel size of the detector, and is approximately 1
.
The exposure time for each dataset was: 30 s for PCH 1, 60 s
for PCH 2a, and 15 s for PCH 2b.
In our analysis, we first applied the standard procedures for correcting and calibrating the SUMER
raw data. They include decompression, reversal, as well as flat-field, dead-time,
local-gain and geometrical corrections. The method used to deduce the line parameters (radiance, central
position of the spectral line and width) is described as follows. We calculated the central position for every pixel, by integrating the line radiance across a certain spectral window and determining
subsequently the location of the 50% level with sub-pixel accuracy. This procedure is
frequently used to obtain SUMER Dopplergrams (see details in Dammasch et al. 1999)
and dramatically reduces the computing time for a large number of data. The results deduced by
this method are statistically consistent with those obtained by using the standard Gaussian fitting
programme (Xia 2003). In addition, a line-position correction was applied, to remove
spurious spectral line shifts caused by thermal deformations of the instrument, and to eliminate
residual errors (systematically varying along the slit) after the geometric correction, using the
standard software (Xia 2003; Dammasch et al. 1999).
![\begin{figure}
\par\includegraphics[width=17cm,clip]{2579fig3.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/Timg15.gif) |
Figure 3:
The evolution of a small region at the solar limb, inside the southern
coronal hole (PCH 2a). The image is a time sequence taken repeatedly every
minute for 14 h. Brighter colors represent higher radiance of
the N IV 765 Å line. |
| Open with DEXTER |
![\begin{figure}
\par\includegraphics[angle=90,width=16.5cm,clip]{2579fig4.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/Timg16.gif) |
Figure 4:
Radiance ratio
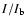 (
(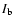 is the background radiance taken from the
darkest pixel at each height) versus time (in min) at 10
and 15
above the limb for PCH 2a (N IV 765 Å line).
The over-plotted curves represent
is the background radiance taken from the
darkest pixel at each height) versus time (in min) at 10
and 15
above the limb for PCH 2a (N IV 765 Å line).
The over-plotted curves represent
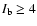 ;
we use them for counting the spicules.
With "A'' and "B'' we denote two locations where spicules show
an obvious periodicity. ;
we use them for counting the spicules.
With "A'' and "B'' we denote two locations where spicules show
an obvious periodicity. |
| Open with DEXTER |
The time evolution of a small region at the solar limb, inside the northern
coronal hole (PCH 1), is displayed in Fig. 2 as radiance maps in the
N IV 765 Å (left) and Ne VIII 770 Å (right) lines.
The maps show a time sequence taken repeatedly every 30 s
for 4 h. In order to increase the visibility of structures, we have
processed this image by using a logarithmic scale to improve its
contrast.
The limb position is set as Y=0. We have determined it as the peak-radiance
position along the solar y-direction in the N IV line. Previous
studies (see Mariska 1992) indicate that the white-light limb would be about 2
lower than this. And it is indeed 2
lower if we look at the continuum in our data.
At the top of the figures, we plot the radiance varying
with time at a height of 10
above the limb.
One finds that spicule structures are clearly seen in the N IV TR line due to the high brightness contrast between spicules
and the dark regions, while they are indistinguishable in the
Ne VIII line. Further inspection of the latter line reveals that its
time-dependent radiance variation above a height of 5
falls in a range
of standard deviation, indicating that the temperature of spicules cannot reach
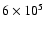 K, the value of the Ne VIII formation temperature. This is
consistent with previous observations (Withbroe 1983; Wilhelm et al. 2000; Budnik et al. 1998).
In the following study, we only use the N IV line due to its strong
radiance and almost pure profile without blends of other lines.
K, the value of the Ne VIII formation temperature. This is
consistent with previous observations (Withbroe 1983; Wilhelm et al. 2000; Budnik et al. 1998).
In the following study, we only use the N IV line due to its strong
radiance and almost pure profile without blends of other lines.
In Fig. 3, we show spicule structures observed in PCH 2a in the
N IV line, with a cadence of one minute, for almost 14 h.
The figure shows solar spicules continuously appearing and then fading
away at the same location, allowing us to see the spicule evolution
with clarity and for a long single observing sequence.
Note that both Figs. 2 and 3 display time-series radiance maps,
which are different than raster images. Spicule shapes in such a map exhibit
their evolution with time if they can be detected at the location where the SUMER
slit is pointed. Therefore, the shapes do not reflect a spatial expansion along
the x-direction. Nevertheless, we are able to see how they expand along the
y-direction (radial direction in these observations) and how long they
persist in the field of view (lifetime). In this context, a narrow (wide)
feature seen in such an image means it has a short (long) lifetime.
In addition, different shapes may represent different evolution stages
of spicules, e.g., in Fig. 3 the expansion front tilts towards
the right side when they are rising and towards the left when falling.
From Figs. 2 and 3, one can see that most spicules have a
height between 10
and 20
(similar in range to a typical
H spicule length). Nevertheless, some of them can reach heights of
30
-40
and can be identified as long macro-spicules
in terms of length.
An exact counting of spicules is difficult due to their different sizes and
overlaps, in particular, for those with small lengths. However, an estimate
can be made for those visible at a height above 10
.
Figure 4
displays the radiance ratio
,
in which
is defined as the background
radiance taken from the darkest pixel at each height, versus time, at 10
and 15
off-limb. The over-plotted curves represent
the radiance ratio larger than 4, and are used to count the number of spicules.
The occurrence rate of spicules during the entire observation, viewed by eye,
is estimated to be about 0.16/min, with little difference between 10
and 15
off-limb.
An average lifetime of about 5 min can be inferred from the occurrence rate
discussed above. An inspection of individual structures indicates that their lifetimes
range from 15 down to 3 min. This is in agreement with previous
studies of H spicules (Beckers 1972), as well as EUV spicules (see, e.g. Budnik et al. 1998; Dere et al. 1989). However, an exact measurement of
the spicule lifetime is not easy, especially for those which are short-lived, because
their roots are overlaid with each other, making them difficult to be determined
individually.
Spicules are sometimes found to have a periodic occurrence, with periods
ranging from 3-5 min to 10-15 min. But as they are so dynamic, they look different along the distance from the limb. For example, one can see in the upper
frame of Fig. 4 at location "A'' a period of 10 min, and at
location "B'' a period of 5 min. Nevertheless, the situation changes
higher up, at 15
:
the structures from location "A'' now show
a finer detail, revealing that each of the three clear peaks from 10
,
"split'' into two, showing a small peak in front of the big one.
This could indicate that such a 10 min-living structure (as seen at 10
above the limb) is actually made up from two independent features,
or that the spicular material is falling back to the surface after rising,
having a slightly different form. Other locations in Fig. 4
also undergo obvious changes in the structure viewed at different heights
off-limb. To further analyse the characteristics of spicules, we look at
the Doppler shift of the plasma in these features.
![\begin{figure}
\par\includegraphics[width=8.8cm,clip]{2579fig5.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/Timg18.gif) |
Figure 5:
Top panel: a segment plot (the first 100 min) taken from PCH 2a showing the
temporal map of the N IV 765 Å radiance, with overlaid contours showing
different levels of radiance in logarithmic scale. The other panels represent
the line Doppler shift (thin black curve) and radiance (the thick grey curve)
varying with time at three locations: at 5
,
10
and 15
above the
limb. Note that the radiance is inverted to enable an easier comparison with the
contours from the top panel. Vertical dotted lines show examples of positions where,
in the third panel (at 10
), the velocity changes its sign (negative values represent
blue-shifts, or outflows; positive values - redshifts, or inflows).
This transition of the spicule Doppler shift could be
caused by material falling after rising. |
| Open with DEXTER |
In Fig. 5, thin black lines in the bottom panels represent
the Doppler velocity for the first 100 min taken from PCH 2a, with the radiance
curve over-plotted as a thick grey line (in arbitrary units). Note that the radiance
is inverted to enable an easier comparison with the contours showing different levels of radiance in logarithmic scale, from the temporal map in the top panel.
We assume that the velocity is zero at the limb, therefore we have subtracted 2 km s-1
from its values. This is, nevertheless, within the limit of the uncertainty for this type
of data, which, in a previous study, we have estimated to be 3 km s-1 (Popescu et al. 2004).
Like the line radiance, the velocities deduced from line shift are highly structured,
with a time scale down to a few minutes. In addition, they seem to vary with height
if observed at the same time.
Obviously, the higher we go off-limb, the more structure we see in spicules,
and the wider become the "inter-spicular'' gaps, with low plasma density.
The precision of the Doppler shift measurement decreases with height,
but not uniformly, as we can still measure it quite well inside the spicules.
To see where the changes in the velocity sign are taking place, we show
some of these examples as vertical dotted lines in Fig. 5; they mark
out the position at 10
above the limb (the third panel) where the Doppler
velocity changes the sign. Following the radiance curves at 10
and 15
off-limb in the one-dimensional plots or in the 2D temporal radiance map (top panel),
we find that the change of sign takes place near the peak of the radiance.
A direct interpretation of such a velocity jump is
that the spicule material is seen falling after rising.
The velocity jump corresponds to locations
where a "falling after rising'' process is also indicated by the expansion
front in the time-series radiance map. Note that the velocity often begins
with a positive sign which means a red shift (i.e. away from the observer).
This can be naturally interpreted as being due to the spicules tilting towards
the back side of the sky plane; in that case the Doppler shift is red when the
material rises and blue when it falls. The opposite situation will happen if
the blue shift is first observed, i.e. the spicules tilt towards the observer.
In order to see in more detail the Doppler velocity behaviour in spicules,
we have chosen another dataset in which the line is cooler (C III 977 Å at 60 000 K) and stronger, so that the exposure time is only 15 s.
Although this dataset lasts only for 15 min, we are able to see a better
time resolution "close-up'' of the Doppler velocity in spicules.
In Fig. 6, we plot the radiance map and Dopplergram of the C III line obtained in PCH 2b, where we find a clear periodic variation of the Doppler shift. The period
is around 5 min, present at all heights between the limb and 15
above it.
Regarding the spicule periodicity, a recent observational and
modeling study exhibited evidence for a relationship between the H spicule
occurrence in an active region and persistent photospheric
oscillation (De Pontieu et al. 2003,2004).
![\begin{figure}
\par\mbox{
\includegraphics[width=8.5cm]{2579fig6a.eps}\includegraphics[width=8.3cm]{2579fig6b.eps} }\end{figure}](/articles/aa/full/2005/30/aa2579-04/Timg19.gif) |
Figure 6:
Time-series Dopplergram and radiance map for the C III 977 Å line,
in PCH 2b, with contours showing levels of radiance in logarithmic scale.
Right panels show the relative Doppler shift at four locations above the limb:
at 0
,
5
,
10
and 15
off-limb. |
| Open with DEXTER |
![\begin{figure}
\par\includegraphics[width=17cm,clip]{2579fig7.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/Timg20.gif) |
Figure 7:
Time-series radiance map and Dopplergram in the N IV 765 Å line of two
long macrospicules taken from PCH 2a. The plotted time intervals are between 460 and 505 min ( left two panels) and 750 and 795 min ( right two panels). The overlaid contours
show radiance in logarithmic scale. |
| Open with DEXTER |
In the presently studied datasets, there are spicules that reach lengths
higher than 30
off-limb (see Figs. 2 and 3).
As discussed in the introduction, we refer to these with the term
"long macro-spicule''. We have looked at the evolution of some of these long
macro-spicules. In Figs. 7 and 8 we plot the time-series radiance
and LOS velocity maps, as derived from the N IV line characteristics, for
the four longest macrospicules from PCH 1 and PCH 2a.
During the observations, the SUMER slit was pointing to the center of the Sun's
poles across the limb. Thus, it practically followed the same region on the Sun for
the whole observing time, with the rotational compensation switched off.
The instrument itself had no mechanical movement between
any two exposures. In addition, the SoHO spacecraft has a good pointing stability better
than 1
over a 15 min interval (Domingo et al. 1995). Therefore, we believe that
the features displayed here were actually from a specific part of a spicule.
When looking at such detailed plots of long macro-spicules one can see how the
radiance of this specific location varies along their lifetime, as well as
the behaviour of the Doppler velocity.
The time-series radiance maps shown in Figs. 7 and 8
reveal that long macro-spicules are highly structured. In the first frame of
Fig. 7, two bright columns with bright feet at the same location near the limb can
be identified between 0
and -10
,
centered at 473 min and 478 min, respectively.
The time interval between them is about 5 min. Another bright column between -5
and -15
centered at 481 min seems to disconnect with the low-lying bright features. We cannot know
whether it is another jet or just the returning plasma of the macro-spicule. A similar
situation can be found at the leading edges of the other long macro-spicule, but less obvious
concerning the 5-min interval for the one centered at 90 min plotted in Fig. 8. For the
macro-spicule centered at 775 min in Fig. 7, individual components from its right
side seem to have different footpoints (a few arcsec away from those on the left).
![\begin{figure}
\par\includegraphics[width=8.8cm,clip]{2579fig8.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/Timg21.gif) |
Figure 8:
The same as Fig. 7, but taken from PCH 1 (the time interval
between 60 and 110 min). |
| Open with DEXTER |
The fact that long macro-spicules are composed of several individual spikes seems to
be a general characteristic for both datasets from PCH 1 and PCH 2a (look again
at Fig. 2, upper panel, left and Fig. 4). Some of them have an obvious
periodicity in the radiance of 5 min, regardless of the exposure time of the dataset.
This corresponds exactly to the average lifetime of a "normal'' spicule, as seen in the same
data. This therefore suggests
that a long macro-spicule is made from a group of spicules that reach larger
heights. It is also possible that two adjacent components could actually represent the
same plasma loop, tilting backward and then towards us (or vice-versa), but this
requires a special configuration: the loop to be oriented along the line
of sight, with both feet inside the slit.
One finds, in the time series Dopplergrams shown in Figs. 7 and 8,
that the amplitude of the Doppler shift variation for the macro-spicules is the
smallest (mostly ranging from -5 to +5 km s-1) just above the limb. The Doppler
shift values quickly increase with height above 10
,
reaching a maximum between 20
and 30
.
The maximum red-shift ranges from 20 to 60 km s-1, while the blue-shift
from -10 to -40 km s-1.
As pointed out before when we discussed Fig. 5, the precision of the Doppler shift
drops together with the plasma density, therefore we can only trust its values inside spicules.
At large off-limb positions outside spicules, the derived velocity values are
not reliable, and thus they have not been plotted.
The two long macro-spicules from Fig. 7 and the left one in Fig. 8 are
found to be composed of several increases in radiance (individual spicules) with
alternating velocity signs. On the contrary, the one centered at 92 min
in Fig. 8 exhibits a different Doppler pattern, without a similar systematical
change of the velocity signs. A possible reason may be that this macro-spicule
was located in the sky plane, so no systematical LOS velocity was detected.
Long macrospicules are often observed having one side with a red-shift and
another side with a blue-shift if seen in the Dopplergrams obtained by CDS/SoHO
(see, e.g. Pike & Mason 1998; Banerjee et al. 2000). This has been interpreted as evidence of rotating
motion of the spicular plasma, which may be driven by torsional Alfvén waves
propagating along an open magnetic flux tube (Kudoh & Shibata 1999). Another possible explanation
is that this would be an indication of a flow up and down along a loop with both feet rooted
on the Sun's surface. In that case, its projection constructs a spike-like structure in
the sky plane.
In our temporal series data, we see that the long macro-spicule consists of several
increases in radiance (individual spicules) with alternating Doppler velocity signs.
This phenomenon could be caused by an up-going followed by a returning
of the macro-spicule's plasma or just simply by individual spicules if they tilted
away from the sky plane.
An interpretation as a flow along a loop seems to require a special geometrical configuration,
as discussed above. Furthermore, we emphasize here that the transition of Doppler
shift actually took place at the same location, on top of the peak. Should a rotating
motion have been considered, we would have had an interesting result that the transverse
velocity changes its sign during the decaying phase of macro-spicules. It is also possible
that, as suggested by Pike & Mason (1998), a combination of both rotating and up-going motion was
indeed observed. Therefore, it seems that at the time being we are unable to get an exclusive
interpretation, and a fast-repeated raster-scanning observation should hopefully provide a
clear view on the time evolution of an entire spicule-structure.
The low velocity in the early stage of spicule evolution deduced from the
line shift has also been found with CDS observations (Pike & Mason 1998; Pike & Harrison 1997).
The reason why spicules have a low Doppler velocity just above the limb is
unclear. One possible explanation is that the measured line profiles are
contributed largely by the background emission there, which stems from
surrounding structures being at rest. From a statistical point of view, velocities from
unresolved spicules with random orientations may also cause a small total shift
by cancellation with each other. Another possibility is that spicules are
really undergoing acceleration at these heights, as suggested by
Pike & Harrison (1997). We believe that a further study is necessary in order to answer this question.
In the radiance map from Fig. 3, many bright
structures show a vertical appearance which might reveal that they are ejected
quickly in this initial stage, an idea consistent with previous studies
(Karovska & Habbal 1994; Parenti et al. 2002).
In this study we analysed the behaviour of solar EUV spicules in three observing
sequences, one of them being as long as 14 h. It is of great interest to
see the spicules continuously appearing and then fading away at the same location,
inside both the northern and the southern polar coronal holes.
Most spicules studied here have a height between 10
and 20
above the
limb. Nevertheless, a few of them can reach a height up to 40
,
being defined then
as "long macro-spicules''. Their lifetime ranges from 15 min down to 3 min.
These results are consistent with previous studies carried out by various
authors cited in the introduction.
We have concluded that EUV spicules occur repeatedly with a birth rate of around 0.16/min
as estimated at 10
above the limb. But their periodicity is difficult to determine,
as spicules have different heights and it is not always possible to distinguish individual features.
Nevertheless, a careful examination of the long datasets in the N IV line, as well as a
short but better resolution data in the C III line indicate that the periods would be
around 5 min.
Another conclusion derived is that the high long-lived macro-spicules
are not a single entity, but a group of plasma jets reaching larger heights, occurring
repeatedly at a time interval similar to the normal spicules, of about 5 min.
A fast-repeated raster-scanning observation is indeed necessary to provide
a clearer view on the time evolution of a spicule, and especially to what produces
the red-blue velocity structure clearly seen in long macro-spicules.
Acknowledgements
Research at Armagh Observatory is grant-aided by the N. Ireland Dept. of Culture,
Arts and Leisure. This work was supported in part by PPARC grant PPA/G/S/2002/00020 and by
the Programme for Research in Irish Third Level Institutions for Grid-enabled Computational
Physics of Natural Phenomena (Cosmogrid). The work of LDX in China was supported by the
Key Program of the Chinese Academy of Science (KZCX3-SW-144). The SUMER project is financially
supported by DLR, CNES, NASA and the ESA PRODEX programme (Swiss contribution). We thank the
EIT team for the full-Sun data.
Finally, we thank the referee for the helpful comments and suggestions.
- Banerjee, D.,
O'Shea, E., & Doyle, J. G. 2000, A&A, 355, 1152 [NASA ADS]
- Beckers,
J. M. 1968, Sol. Phys., 3, 367 [NASA ADS]
- Beckers,
J. M. 1972, ARA&A, 10, 73 [NASA ADS]
- Bohlin,
J. D., Vogel, S. N., Purcell, J. D., et al.
1975, ApJ, 197, L133 [NASA ADS] [CrossRef] (In the text)
- Budnik, F.,
Schröder, K.-P., Wilhelm, K., & Glassmeier, K.-H. 1998,
A&A, 334, L77 [NASA ADS]
- Dammasch,
I. E., Wilhelm, K., Curdt, W., & Hassler, D. M. 1999,
A&A, 346, 285 [NASA ADS] (In the text)
- De Pontieu,
B., Erdélyi, R., & de Wijn, A. G. 2003, ApJ, 595,
L63 [NASA ADS] [CrossRef]
- De Pontieu,
B., Erdélyi, R., & James, S. P. 2004, Nature, 430,
536 [NASA ADS] [CrossRef]
- Dere, K. P.,
Bartoe, J.-D. F., & Brueckner, G. E. 1983, ApJ, 267,
L65 [NASA ADS] [CrossRef]
- Dere, K. P.,
Bartoe, J.-D. F., Brueckner, G. E., & Recely, F.
1989, ApJ, 345, L95 [NASA ADS] [CrossRef]
- Domingo, V.,
Fleck, B., & Poland, A. I. 1995, Sol. Phys., 162, 1 [NASA ADS] [CrossRef] (In the text)
- Karovska, M.,
& Habbal, S. 1994, ApJ, 431, L59 [NASA ADS] [CrossRef]
- Kudoh, T., &
Shibata, K. 1999, ApJ, 514, 493 [NASA ADS] [CrossRef] (In the text)
- Lemaire, P.,
Wilhelm, K., Curdt, W., et al. 1997, Sol. Phys., 170, 105 [NASA ADS] [CrossRef]
- Mariska,
J. T. 1992, Cambridge Astrophys. Ser., The Solar Transition
Region (Cambridge Univ. Press), 23
(In the text)
- Moore, R. L.,
Tang, F., Bohlin, J., & Golub, L. 1977, ApJ, 218, 286 [NASA ADS] [CrossRef] (In the text)
- Parenti, S.,
Bromage, B., & Bromage, G. E. 2002, A&A, 384, 303 [EDP Sciences] [NASA ADS] [CrossRef]
- Pike, C., &
Harrison, R. 1997, Sol. Phys., 175, 457 [NASA ADS] [CrossRef]
- Pike, C., & Mason,
H. 1998, Sol. Phys., 182, 333 [NASA ADS] [CrossRef]
- Popescu,
M. D., Doyle, J. G., & Xia, L. D. 2004, A&A,
421, 339 [EDP Sciences] [NASA ADS] [CrossRef] (In the text)
- Roberts,
W. O. 1945, ApJ, 101, 136 [NASA ADS] [CrossRef]
- Secchi, A.
1877, Le Soleil (Paris: Gauthier-Villars), 2
- Sterling,
A. C. 2000, Sol. Phys., 196, 79 [NASA ADS] [CrossRef] (In the text)
- Wilhelm, K.
2000, A&A, 360, 351 [NASA ADS]
- Wilhelm, K.,
Curdt, W., Marsch, E., et al. 1995, Sol. Phys., 162, 189 [NASA ADS] [CrossRef]
- Wilhelm, K.,
Lemaire, P., Curdt, W., et al. 1997, Sol. Phys., 170, 75 [NASA ADS] [CrossRef]
- Wilhelm, K.,
Dammasch, I. E., Marsch, E., & Hassler, D. M. 2000,
A&A, 353, 749 [NASA ADS]
- Withbroe,
G. L. 1983, ApJ, 267, 825 [NASA ADS] [CrossRef]
- Xia, L. D. 2003,
Ph.D. Thesis, Georg-August-Universität, Göttingen
(In the text)
Copyright ESO 2005
![\begin{figure}
\par\includegraphics[width=8.8cm,clip]{2579fig1.ps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/img13.gif)
![\begin{figure}
\par\includegraphics[width=16.8cm,clip]{2579fig2.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/img14.gif)
![\begin{figure}
\par\includegraphics[width=17cm,clip]{2579fig3.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/img15.gif)
![\begin{figure}
\par\includegraphics[angle=90,width=16.5cm,clip]{2579fig4.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/img16.gif)
![\begin{figure}
\par\includegraphics[width=8.8cm,clip]{2579fig5.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/img18.gif)
![\begin{figure}
\par\mbox{
\includegraphics[width=8.5cm]{2579fig6a.eps}\includegraphics[width=8.3cm]{2579fig6b.eps} }\end{figure}](/articles/aa/full/2005/30/aa2579-04/img19.gif)
![\begin{figure}
\par\includegraphics[width=17cm,clip]{2579fig7.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/img20.gif)
![\begin{figure}
\par\includegraphics[width=8.8cm,clip]{2579fig8.eps}
\end{figure}](/articles/aa/full/2005/30/aa2579-04/img21.gif)